|
| |||
|
Týdeník věnovaný aktualitám a novinkám z fyziky a astronomie. | |||
|
Laboratoř na kabelech
Petr Panchártek
Již od pradávna tu žije společně s námi létající hmyz. Hmyzí technika létání se měnila společně s prostředím, ve kterém hmyz žil. Přesto je experimentální studium velmi obtížné z důvodu jejich malé velikosti a vysoké rychlosti pohybu. Existuje však další možnost, robot, který je řízený pomocí kabelů a jmenuje se „Laboratoř na kabelech“. Tento robot slouží pro sledování volně létajícího hmyzu a interakci s ním. Na kabelech jsou namontované sledovací kamery, které se pohybují automaticky s hmyzem. Skupina vědců z francouzského CNRS navrhla rychle reagující kontrolér, který minimalizuje chybu online sledování mezi pozicí létajícího hmyzu, kterou poskytuje vestavěný stereosystém, a polohou pohybující se laboratoře vypočtené z délek kabelů. „Laboratoř na kabelech“ byla testována při pozorování můry osenice ypsilonové (dlouhé asi dva centimetry), která létala rychlostí až tři metry za sekundu. S pomocí předem zaznamenaných trajektorií lze sledovat i jiný hmyz, jako jsou ovocné mušky nebo komáři. „Laboratoř na kabelech“ je významná jak pro studie volného letu, tak letu ovlivněného feromony či jinými stimuly, kdy dochází k modulaci letového chování (například za větru, tehdy hovoříme o tzv. anemotaxi).

Robot „Laboratoř na kabelech“. Zdroj: CNRS.
Počáteční experimenty
Hmyz má sice miniaturní mozek, ale jeho reflexy nejsou vůbec jednoduché. Naopak umožňují velmi bohaté reakce na vnější podněty. Například octomilka obecná je malý hmyz vážící půl miligramu, který má pouze sto tisíc neuronů (milionkrát méně než lidský mozek). Ani takto snížený počet neuronů neomezí smyslové zpracovávání techniky letu a letové manévry, které jsou nesrovnatelné se současnými letovými technologiemi. Jako zdroj inspirace pro budoucí vývoj robotiky by mohlo sloužit pochopení, jak miniaturní hmyzí mozky ovládají smyslové zpracování a chování za letu. Představte si například mikrovzdušná vozidla napodobující mávání letu v hmyzím měřítku nebo čichové roboty inspirované anemotaxií můr.
Vědci po celá desetiletí vyvíjejí laboratorní experimentální sestavy pro studium a pochopení letového chování hmyzu. Existují dva principy, jak studovat letovou techniku hmyzu. První princip spočívá v udržení hmyzu v jedné poloze, v níž se letová kinematika podrobně analyzuje vysokorychlostními kamerami. Druhý princip bere v úvahu přirozenější podmínky, tj. volný let.
Omezení pohybu zvířat je možné uskutečnit dvěmy způsoby. První z možností využívá hmyzu se specifickými vlastnostmi, například lišajové jsou schopni se při krmení nektarem vznášet před umělými květinami (viz obrázek A níže). Druhou možností je fyzické omezení hmyzu – například postrojem (viz obrázek C níže). Ani jedna z možností není nejlepší. Buď jsme přinuceni jen ke studiu ustáleného stavu, tj. vznášení za letu, nebo využíváme tuhý postroj, který ovlivňuje letovou dynamiku, protože neumožnuje pohyby při naklápění nebo stoupání.

Experimentální sestavy pro výzkum létajícího hmyzu. (A) Manduca sexta (rozpětí křídel deset centimetrů) vznášející se před umělou květinou. (B) Letecká aréna, virtuální realita. (C) Přivázaná můra (Osenice ypsilonová). (D) „Laboratoř na kabelech“ popisovaná v tomto bulletinu. Zdroj: Science Robotics.
Jiný přístup
Při studiu principů volného letu se vědci nejprve pokusili zaznamenat svalovou aktivitu pomocí miniaturních elektrofyziologických zařízení, které nesl hmyz během letu. Tento způsob měl jen částečný úspěch, protože nesené zatížení ovlivňuje výkon letu, a je tedy vhodné jen pro největší hmyz (například lišaje s tělesnou hmotností od jednoho do tří gramů), který má schopnost přenášet zátěž až do deseti procent své tělesné hmotnosti.
Dále byly využívány větrné tunely (cca 1 až 2 m dlouhé a široké 30 cm), které byly vybavené zařízením pro zachycování pohybu, což je běžný nástroj pro zaznamenávání trajektorií hmyzu při volném letu. Některé experimenty zkoušely detekovat přitažlivost hmyzu v turbulentních pachových oblacích, jako je anemotaxie samců můr na sexuální feromon. Letové cesty jsou obvykle klikaté podél osy větru s amplitudou, která se snižuje, když se hmyz blíží ke zdroji. Přesto se jedná pouze o průměrné nebo nepřímé pozorování, protože větrné tunely neumožňují přesné dodání podnětů v prostoru a čase. Externí kamery jsou v turbulentním tunelu umístěné daleko od hmyzu, a to brání jakékoli analýze letového chování. To je jedním z důvodů, proč je obtížné dosáhnout synchronního měření podnětů ve vzduchu, s nimiž se zvíře skutečně setkává v kterémkoliv bodě letu v závislosti na orientaci směrem ke zdroji.
Naprostá většina experimentů zkoumajících let hmyzu se tedy omezila jen na studium vznášejících se letů (viz obrázek A výše), uvázaných letů (viz obrázek C výše) nebo omezených letů ve stísněném prostředí (viz obrázek B výše). Vznikl však robot, který se jmenuje „Laboratoř na kabelech“ a který se snaží tato omezení překonat. Laboratorní zařízení je připojené na kabelech, takže se může pohybovat spolu s pozorovaným zvířetem (viz obrázek D výše). Kabeloví roboti patří do zvláštní třídy paralelních robotů, u nichž jsou pevné vodiče nahrazeny ohebnými kabely. Nabízejí několik výhod, jako je například možnost pohybovat objekty s vysokou přesností ve velkém pracovním prostoru. Velmi populární aplikace jsou SpiderCam nebo SkyCam – zavěšené kamery pohybující se na stadionech – nebo robotický jeřáb RoboCrane používaný na staveništích. Využití laboratorních souprav pohybujících se spolu s létajícím hmyzem je náročné, protože trajektorie hmyzu není předem známa a rychlost letu může být vysoká (až několik metrů za sekundu).

Kamera Spidercam LIGHT byla navržena pro práci na menších místech,
jako jsou televizní studia nebo sportovní haly. Zdroj: SpiderCam.
Laboratoř na kabelech
Tento robot má celkem šest stupňů volnosti. Sleduje létající hmyz v pracovním prostoru o délce 6 m, šířce 4 m a výšce 3 m (viz video a obrázek A níže). Koncový prvek, nazývaný létající rám, je otevřenou krychlí s délkou hrany 30 cm, ve které může hmyz volně létat. Létající rám je uchycen na kabelech a při sledování dráhy létajícího hmyzu je ovládaný motorovými navijáky (viz obrázek B a C níže). Do létajícího rámu byl integrován optický systém, který vypočítává online trojrozměrnou (3D) polohu XT hmyzu pomocí infračerveného záření a kalibrovaných kamer. Strategie řízení, která spočívá v pronásledování aktuální cílové polohy, je riskantní a může vést mimo trať, protože hmyz může létat relativně vysokou rychlostí. Místo toho bere řídicí schéma v úvahu směr pohybu hmyzu a předpovídá, jaké bude jeho budoucí umístění a směřuje robota k němu (viz obrázek D níže). Na tuto strategii lze pohlížet jako na druh odkloněného pronásledování, které se také používá při navádění raket. Jedním z hlavních rozdílů je však to, že raketa zaměřená na zachycení cíle letí svou maximální rychlostí, zatímco laboratoř na kabelech sleduje rychlost hmyzu nepřetržitě. Rychlost sledování lze popsat rovnicí (X je pozice středu létajícího rámu, XT je poloha hmyzu, VT je rychlost hmyzu, Kp a Kd jsou nastavitelné koeficienty)
V = Kp(XT − X) + Kd VT.
Dlouhodobá expozice „Laboratoře na kabelech“ při rychlosti 1 m/s.
Zdroj:
Science Robotics.
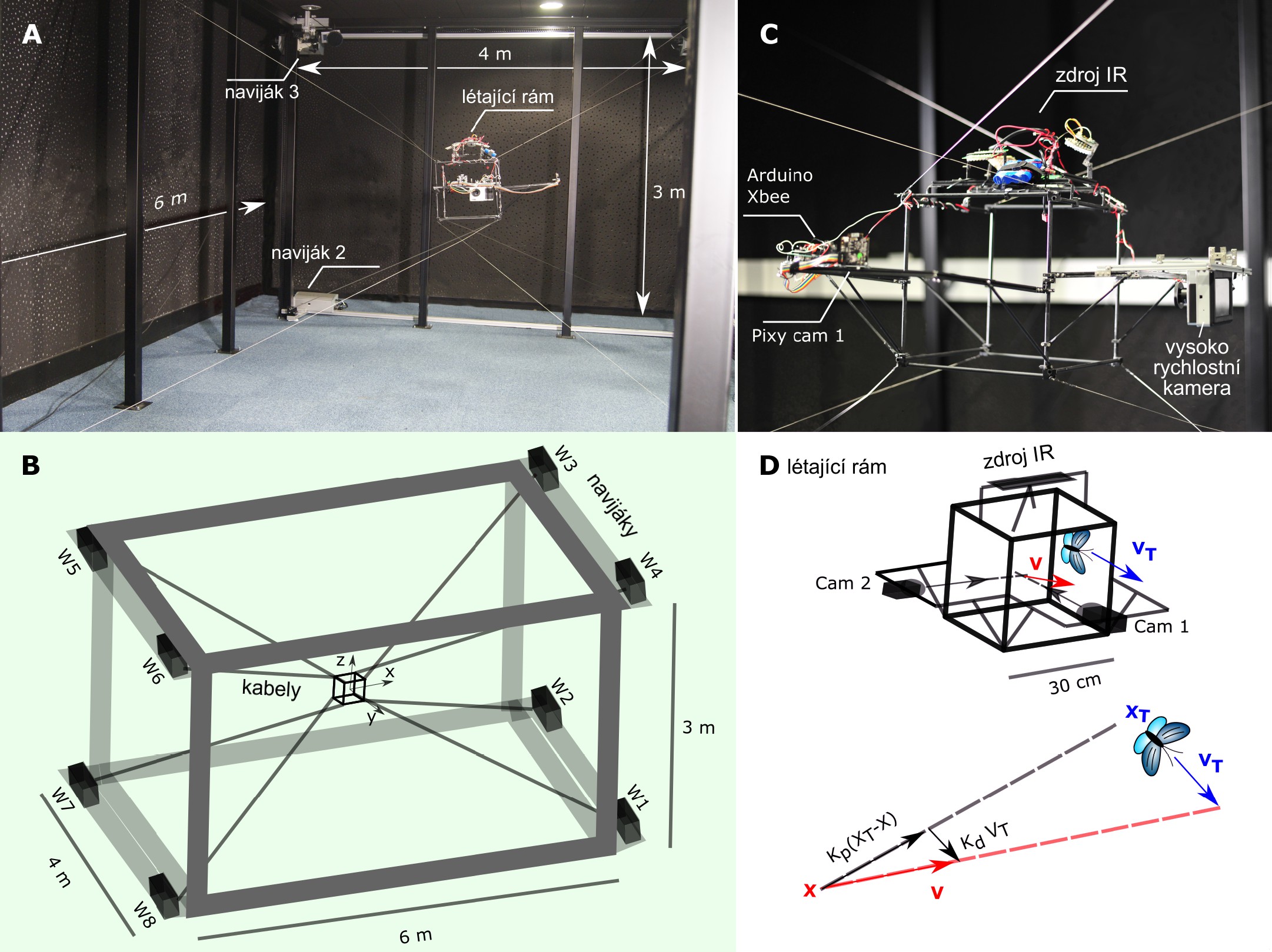
Nastavení „Laboratoře na kabelech“. (A) Fotografie kabelového robota (6×4×3 m). (B) Schematické zobrazení sestavy. (C) Fotografie létajícího rámu (30×30×30 cm). Létající rám s sebou nese laboratorní vybavení, tj. zdroj infračerveného záření a dvojici kalibrovaných kamer (Pixy cam 1 a 2) pro online sledování pozice hmyzu. Létající rám se automaticky pohybuje, aby udržel hmyz v detekčním dosahu kamer. (D) Řízení robota jako odchýlené pronásledování. Umístění robota a hmyzu jsou X a XT. Rychlost hmyzu je označena VT. Rychlost sledování V je součtem „pronásledovacího“ členu směřujícího od robota k objektu a korekčního členu, který bere v úvahu směr pohybu cíle. Zdroj: Science Robotics.
Experimentální ověření návrhu řízení
Nejprve byly provedeny robotické experimenty s trajektoriemi hmyzu, které byly již dříve zaznamenány ve větrných tunelech. Trajektorie odpovídaly různým druhům hmyzu, který létal v pro hmyz zajímavých pachových oblacích:
| druh hmyzu | počet jedinců | oblak |
|---|---|---|
| octomilka obecná (ovocná muška) | 169 | etanol |
| komár tropický (komár) | 65 | CO2 |
| osenice ypsilonová (můra) | 38 | feromon |
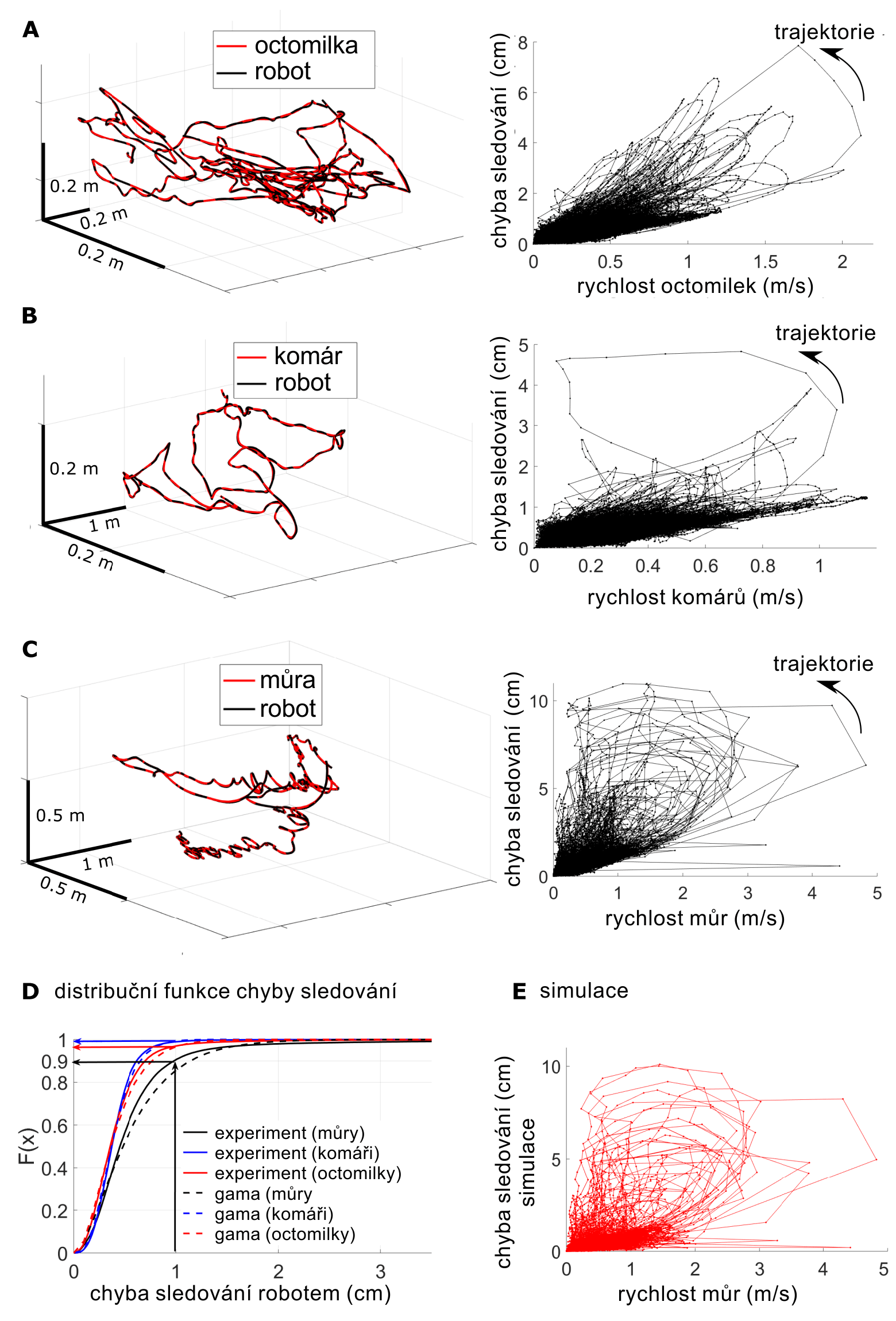
Sledování předem zaznamenaných trajektorií hmyzu. (A, B, C) nalevo: příklady trajektorií vybraného hmyzu (červené křivky) oproti trajektorii robota (černé křivky). (A, B, C) napravo: chyby sledování robota v závislosti na rychlosti letu hmyzu. (D) kumulativní rozdělení chyby sledování (tzv. distribuční funkce) pro experimentální data (jednoduché křivky) a teoretické gama rozdělení (přerušované křivky). (E) chyby sledování simulovaného modelu a rychlost hmyzu (údaje o můrách). Zdroj: Science Robotics.
Simulace předem zaznamenaných trajektorií byla zaměřena na dva cíle: 1) provádění předběžných zkoušek s různými druhy hmyzu; 2) obcházení vestavěného detekčního systému, aby došlo k ověření návrhu robota. Na předchozím obrázku jsou tři příklady (A až C) trajektorií sledovaných robotem (řídicí koeficienty Kp = 8,4 a Kd = 1). Robot byl naprogramován tak, aby každých deset milisekund dosáhl nové cílové polohy, která byla postupně čtena z datového souboru předem zaznamenané trajektorie. Pro test účinnosti robota se nejprve vypočítala chyba sledování pro všechny body ve všech trajektoriích jako euklidovská vzdálenost mezi cílem (létající hmyz) a středem létajícího rámu. Chyba sledování se zvyšovala s rychlostí hmyzu, takže pro nízké rychlosti byly zaznamenány malé chyby, a naopak pro vysoké rychlosti velké chyby (viz obrázky A až C napravo). U některých trajektorií vznikaly velké zbytkové chyby, ke kterým došlo při vysoké rychlosti. Přetrvávaly od jednoho provozního cyklu do dalšího (tj. 10 ms), i když hmyz zpomalil. Toto pozorování naznačuje určitá omezení robota, pokud jde o rychlost a zrychlení. K posouzení těchto hranic byla analyzována přechodná odezva robota v závislosti na vstupech pro řízení rychlosti. Ukázalo se, že robot má hranici použitelnosti při rychlosti 3,6 m/s a zrychlení 17 m/s2. Kumulativní rozdělení znázorněné na obrázku D pro různé druhy hmyzu naznačuje, že ve více než 90 % případů byla chyba sledování menší než 1 cm, což je ve srovnání s délkou létajícího rámu (30 cm) málo.
Analýza letové kinematiky
Důležitou součástí při sledování létajícího hmyzu je i způsob jeho vzletu (viz obrázek A níže). Hmyz se jemně umístí na vzletovou plošinu (2 cm2), která je blízko středu létajícího rámu a robot začne okamžitě ve sledování. Pokud ke spontánnímu vzletu nedojde do pěti minut, je let stimulován tepelně (Peltierovým článkem na vzletové plošině). Po vzletu se aktivuje elektromagnet, který přitáhne plošinu, aby nebránil sledování hmyzu robotem. Nebyl zaznamenán žádný rozdíl mezi spontánními vzlety a těmi, které vyvolala tepelná stimulace. V obou případech iniciace letu vzlétla můra osenice ypsilonová tak, že vyskočila z plošiny pomocí křídel a nohou. Takové zahájení letu je pro můry typické.
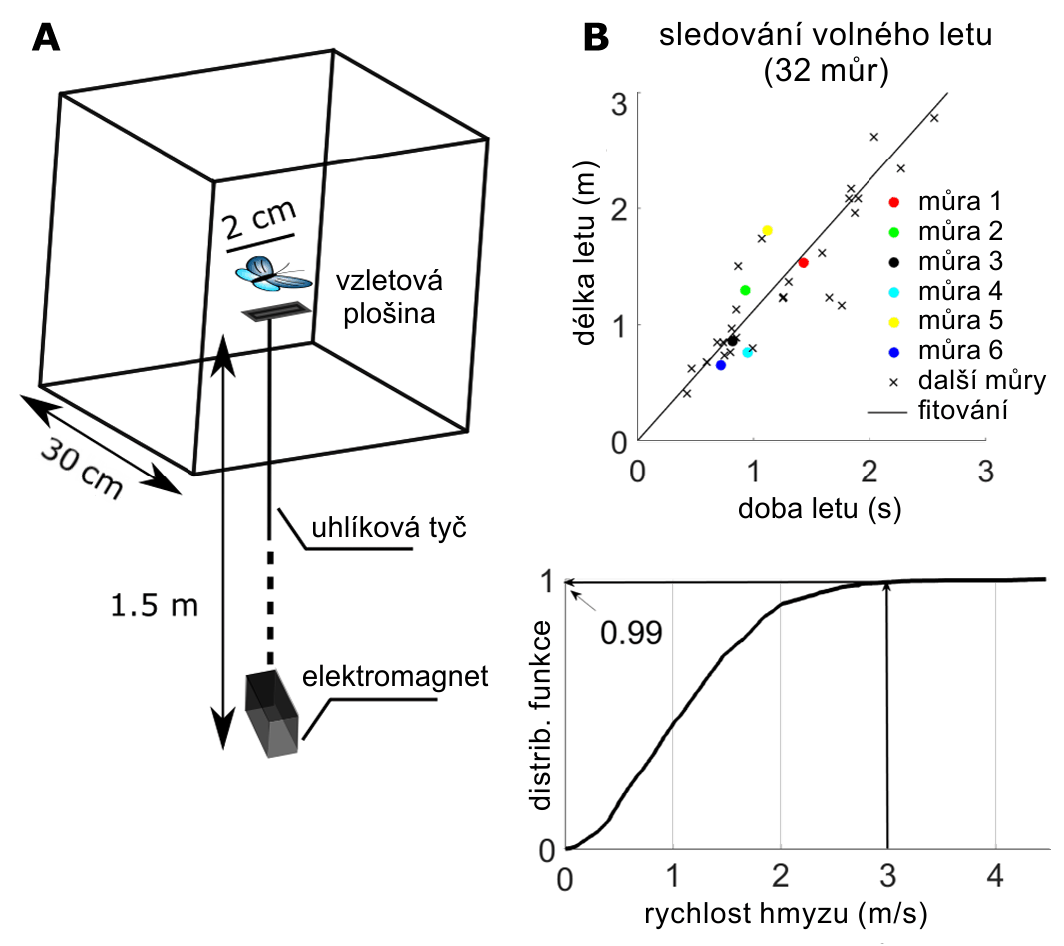
Sledování můr osenice ypsilonové. (A) postup vzletu. (B) sledování volných letů (32 můr). Nahoře: délka letu v závislosti na době letu (lineární regrese). Barevné body znázorňují šest můr použitých při analýze letové kinematiky. Dole: kumulativní pravděpodobnost (distribuční funkce) rychlosti letu hmyzu. Zdroj: Science Robotics.
Experimenty byly prováděny s 32 můrami. Během letu byl hmyz lokalizován online z integrovaného optického systému, a proto byla na rozdíl od předchozích experimentů poloha cíle (hmyzu) zašuměná. Pro zlepšení stability v přítomnosti šumu experimentátoři snížili řídicí koeficienty na hodnoty Kp = 3 a Kd = 0,9. Letové dráhy se pohybovaly od 0,5 do 3 m (viz obrázek B výše). Kumulativní rozdělovací funkce na obrázku B ukazuje, že rychlost hmyzu během letu je v 99 % případů nižší než 3 m/s.
Pro šest můr (znázorněných jako barevné body na obrázku výše) byl zaznamenáván let rychlostí 400 snímků za sekundu, tj. s desetkrát vyšší hodnotou, než je frekvence křídel můry osenice ypsilonové. Vysokorychlostní kamera byla namontovaná na létajícím rámu (viz obrázek A níže). Tělesná kinematika osenice ypsilonové byla na základě nahrané videosekvence analyzována ve 3D počítačovém grafickém programu Blender (obrázek A vpravo). Na základě této analýzy byl udělán počítačový model pohybu, který odděleně popisuje polohu, rotaci těla a pohyb křídel. Lze nastavit i omezení symetrie mezi levým a pravým křídlem. Poloha, rotace těla a pohyb křídel byly optimalizovány poloautomaticky tak, aby odpovídaly pohybu skutečného hmyzu v každém obrazu zaznamenaných videosekvencí. Z přizpůsobeného modelu se extrahovala kinematika těla představovaná náklonem těla (α) a úhlem roviny zdvihu křídla vzhledem k vodorovné rovině (ß), jak je znázorněno na obrázku A (vpravo). Rovina zdvihu křídla byla vypočtena metodou nejmenších čtverců z poloh základny a špičky křídla během celého zdvihu křídla (z přibližně deseti po sobě jdoucích 3D bodů).
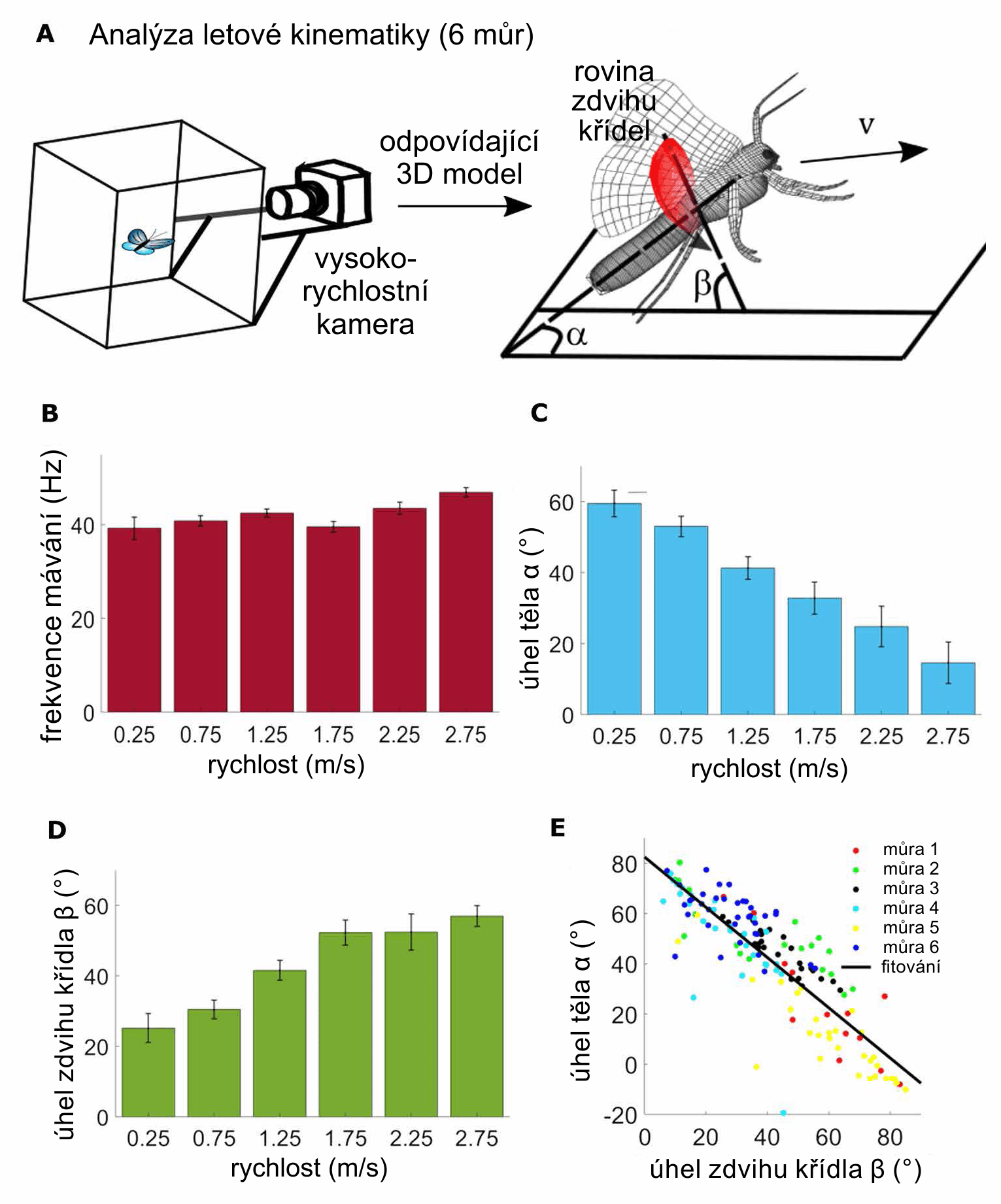
Analýza letové kinematiky. (A) Vysokorychlostní kamera je namontována na létajícím rámu a 3D pozice modelu můry je optimalizována poloautomaticky tak, aby odpovídala skutečnému hmyzu ze zaznamenaných videí. Sklon těla je popsán úhlem ?, tj. úhlem mezi podélnou osou těla a vodorovnou rovinou. ß je úhel mezi vodorovnou rovinou a rovinou zdvihu křídla definovanou polohami základny a špičky křídla během celého zdvihu křídla. (B) Závislost frekvence mávání křídel na rychlosti letu. (C) Závislost úhlu těla na rychlost letu. (D) Závislost úhlu zdvihu křídla na rychlosti letu. V (B) až (D) je průměr v rámci rychlostních intervalů (velikost intervalu 0,5 m/s). (E) Závislost úhlu těla na úhlu zdvihu křídla. Zdroj: Science Robotics.
Sledování můry osenice ypsilonové, extrakce letové kinematiky a porovnávání založené na 3D modelu (skutečná můra v bílé barvě a 3D model v růžové barvě). Zdroj: Science Robotics.
Závěr
„Laboratoř na kabelech“ sledovala volný let můry osenice ypsilonové až do rychlosti 3 m/s. Kinematika můry byla zaznamenávána vysokorychlostní kamerou. Analýza dat ukazuje, že při vyšších rychlostech osenice ypsilonová nakloní své tělo směrem dolů, zatímco rovina zdvihu se stává svislejší. „Laboratoř na kabelech“ se stala důležitým nástrojem pro studium létací techniky hmyzu. Využívá výhody kabelových robotů ke sledování létajícího hmyzu a umožňuje pozorování velmi rychlé dynamiky s malým odporem vzduchu. Díky lehké konstrukci kabelů je možné nasazení ve velkém pracovním prostoru. „Laboratoř na kabelech“ je vhodná pro zaznamenávání volného pohyb hmyzu a je omezena jen velikostí pracovního prostoru. Toto omezení lze změnit délkou kabelů a změnou mechanických navijáků. Experimentovat bude proto možné i ve větším měřítku, a dokonce i v přírodě.
* * *
„Podívej se hluboko do přírody, a pak všechno lépe pochopíš.“
Albert Einstein
Odkazy
![]()
![]()
